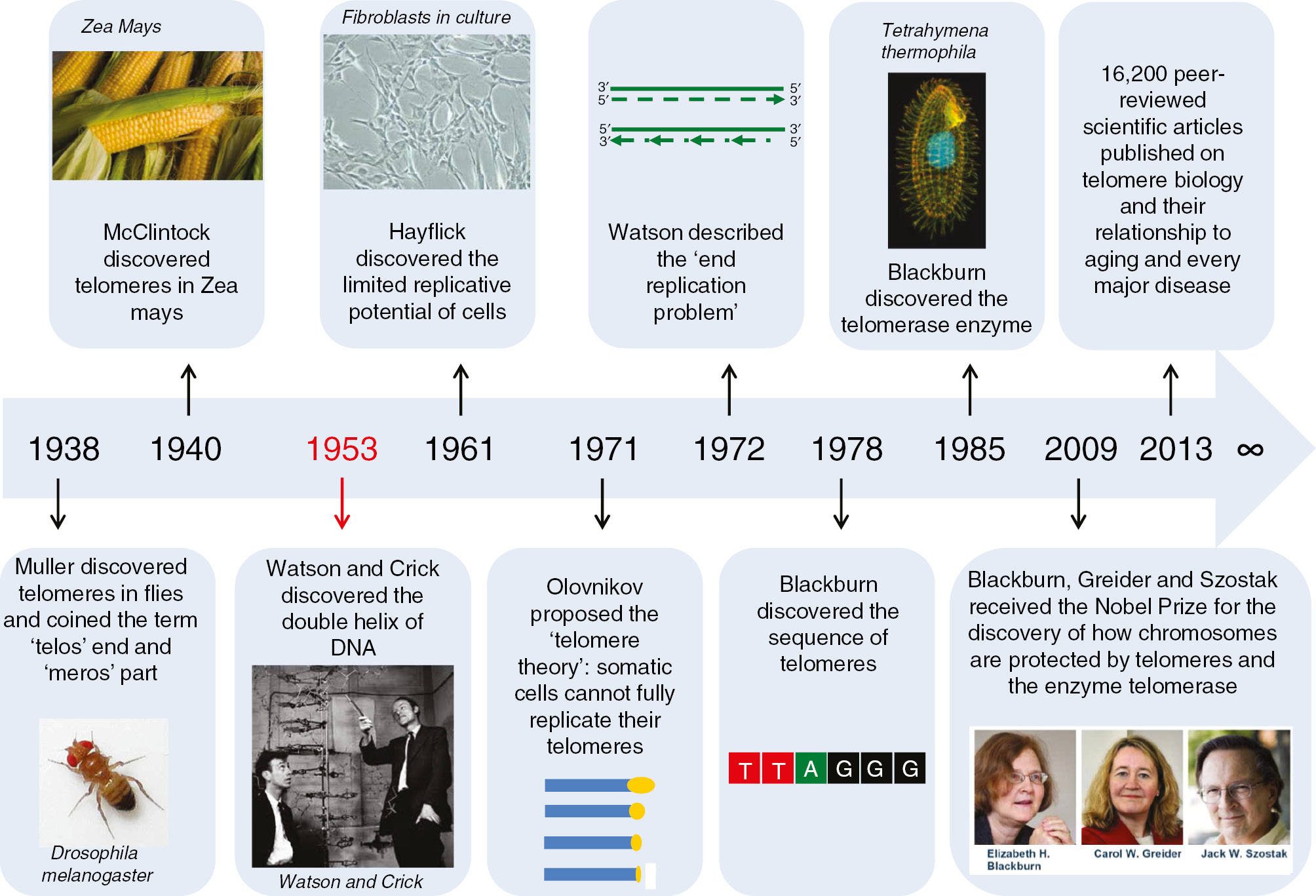
Telomer (“telos-meros”) τέλος ve μέρος sözcüklerinden türetildi (telos “son,” ve meros “parça” demektir).
Telomerler her bir kromozom kolunun ucunu kapayan ve genom sağlamlığını sürdürme işlevini gören nükleoprotein yapılarıdır.
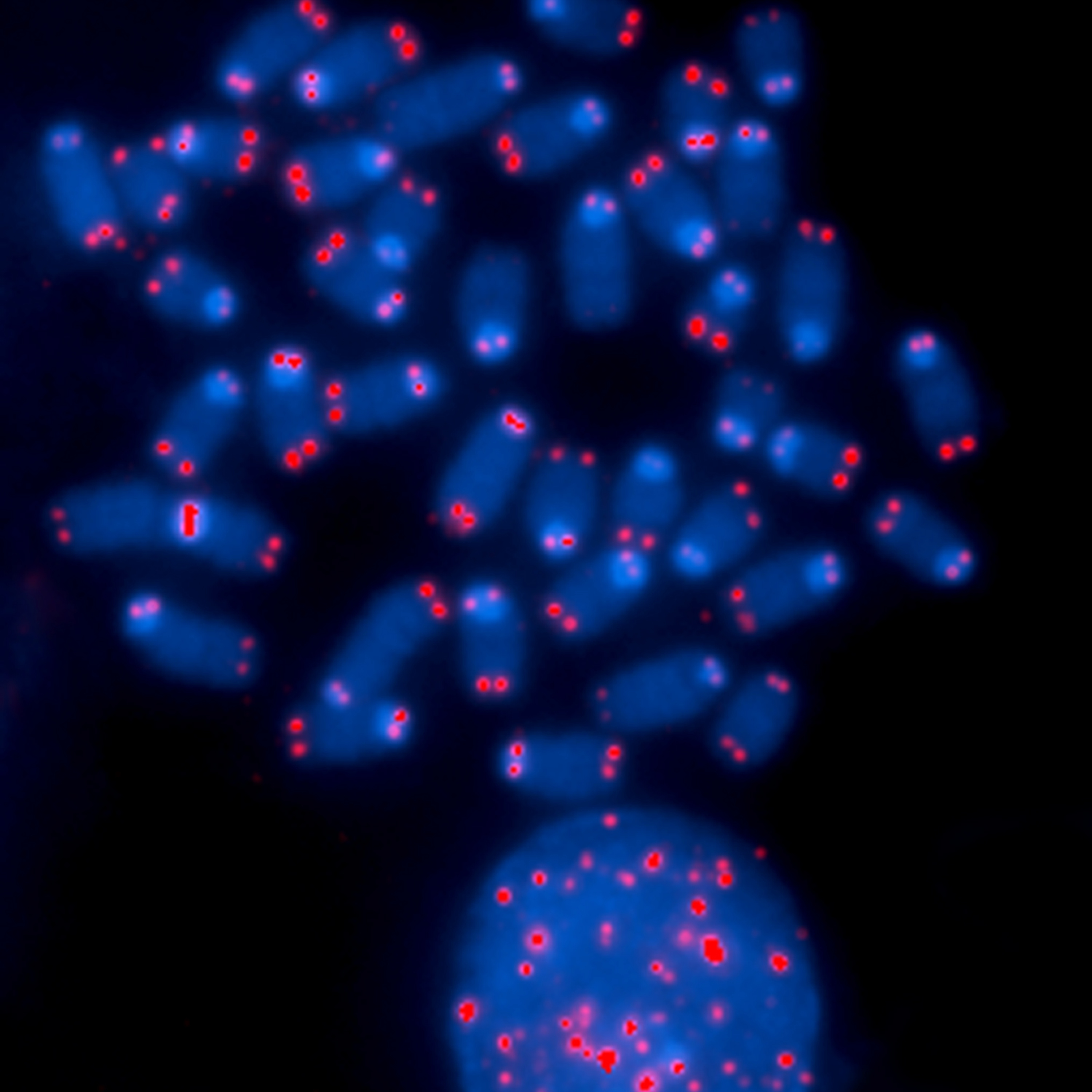
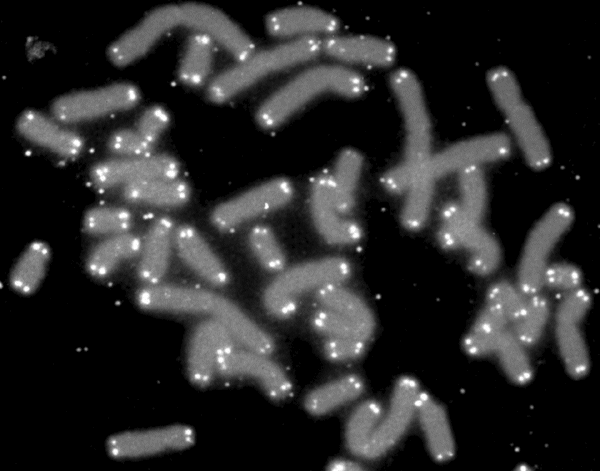
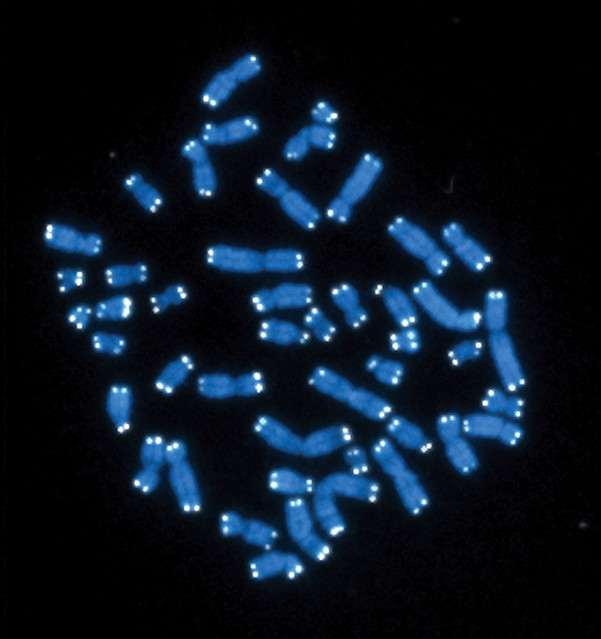
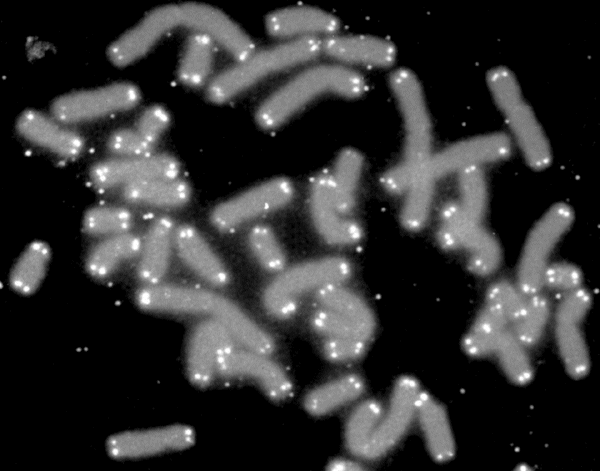
Telomerler (beyaz) ile keplenen insan kromozomları (gri).
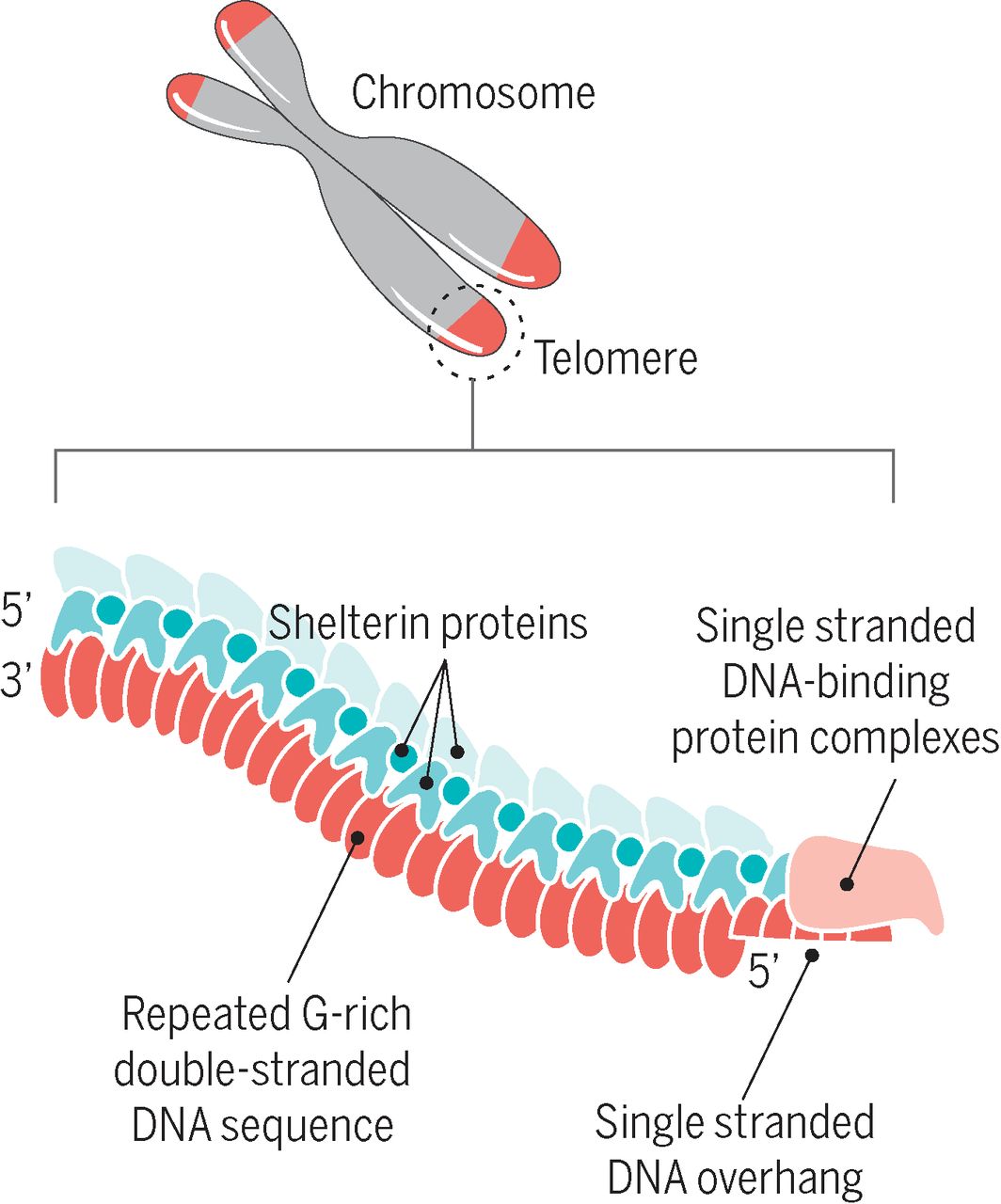
Telomerler doğrusal kromozomların uçlarına yerleşmiş ve kodlama yapmayan yineleyici DNA dizilerinden ve Shelterin karmaşasını oluşturan ek proteinlerden oluşur.
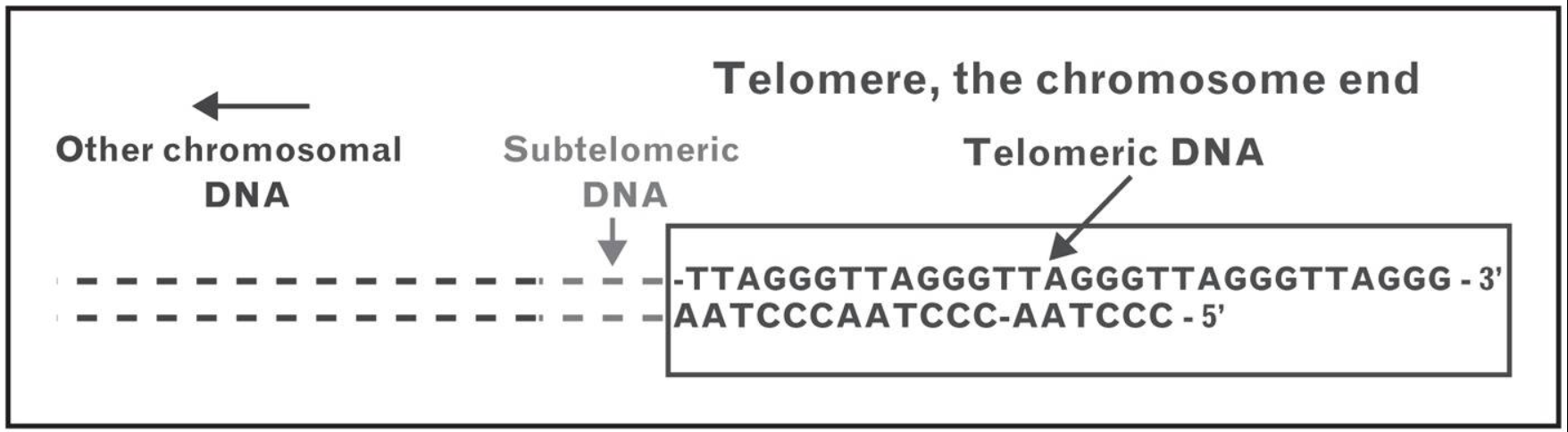
İnsanlarda Omurgalılar için telomerlerdeki nükleotid dizisi 5′-TTAGGG-3′, tümleyici DNA teli 3′-AATCCC-5′ biçimindedir, ve tek-telli TTAGGG uzantı olarak bulunur.
TTAGGG dizisi insanlarda yaklaşık 2.500 kez yinelenir.
İnsanlarda, ortalama telomer uzunluğu doğumda 11 kilobazdan yaşlılıkta 4 kilobaz altına düşer.
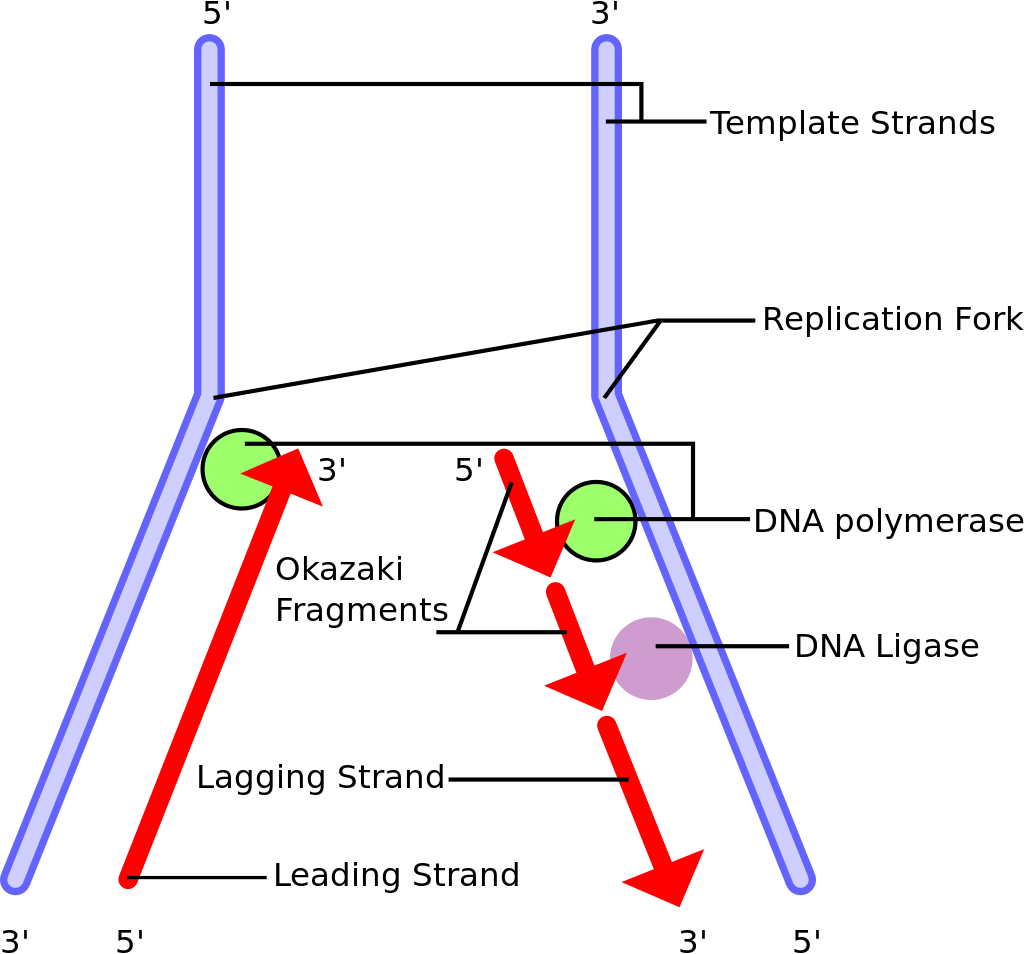
Kromozom eşlenimi sırasında DNAyı eşlemleyen enzimler eşlemlemeyi bir kromozomun sonuna dek sürdüremezler.
Bu nedenle her kromozom eşlenimi sırasında kromozomun ucu kısalır (çünkü Okozaki fragmanlarının bireşimi geciken tele RNA primerlerin önden eklenmesini gerektirir).
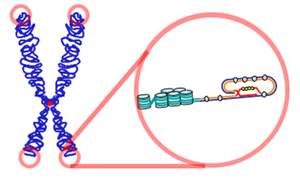
T-ilmeği ve D-ilmeği. G-varsıl telin (mavi) 3′ ucu telomerin tek telli bir uzantısı olarak öne çıkar. Bu G-teli çıkıntısı bir T-ilmeği oluşturmak üzere geriye doğru döner ve 5′ çift-telli telomerik sarmal üzerine uzanarak bir D-ilmeği oluşturur. (LINK)
T-ilmeği ve D-ilmeği. G varsıl telin (mavi) 3′ ucu telomerin tek telli bir uzantısı olarak öne çıkar. Bu G-teli çıkıntısı bir T-ilmeği oluşturmak üzere geriye doğru döner ve 5′ çift-telli telomerik sarmalın üzerine yatarak bir D-ilmeği oluşturur. (LINK)
İlmekli yapı G-varsıl tek-telli bir çıkıntı üretecek bir yolda telomerik DNAnın uçlarında nükleotik etkinlik tarafından oluşturulur.
Bu ilmek geriye dönerek çift-telli telomerik zincirin üzerine yatar ve gevşek DNA uçlarının içsel olarak nükleoprotein yapının içerisine yerleşmesini sağlar.
Prokaryotların çoğunda kromozomlar dairesel olduğu için telomerler bulunmaz.
Ökaryotlarda, telomerler kromozom uçlarında tam-olmayan yarı-tutucu DNA eşleniminin yarattığı sorunu çözer.
Shelterin olarak bilinen bir protein karmaşası telomerlerin uçlarının çift-tel kopmaları (L) olarak tanınmasının önüne geçer (ve bunu homolog yeniden-bileşimi (HR) ve homolog-olmayan uç birleşmesini (NHEJ) engelleyerek yapar).
Doğrusal DNA moleküllerinin uçlarının kısalması. Bu çizgede DNA molekülünün bir telinin ucu iki eşlenim adımı boyunca izlenmektedir. Birinci adımdan sonra, yeni geciken tel kalıbından daha kısadır. İkinci adımdan sonra, hem öncü hem de geciken teller başlangıçtaki anne DNAdan kısa kalmıştır. Bu DNA moleküllerinin burada gösterilmeyen öteki uçları da kısalmıştır.
Ökaryotik DNA eşlenim enzimleri (DNA polimeraz protein karmaşası) kromozomların uçlarında bulunan dizileri (kromatid lifleri) eşlemleyemez ve bu nedenle bu diziler ve taşıdıkları bilgiler yitebilir.
Telomerler uç dizilere “kep” takar ve kendileri DNA eşlenim sürecinde yiter.
Hücrede telomeraz enzimi yineleyici nükleotid dizilerini DNAnın uçlarına ekleme görevini yerine getirir.
Telomeraz telomer “kep”ini yeniden doldurur.
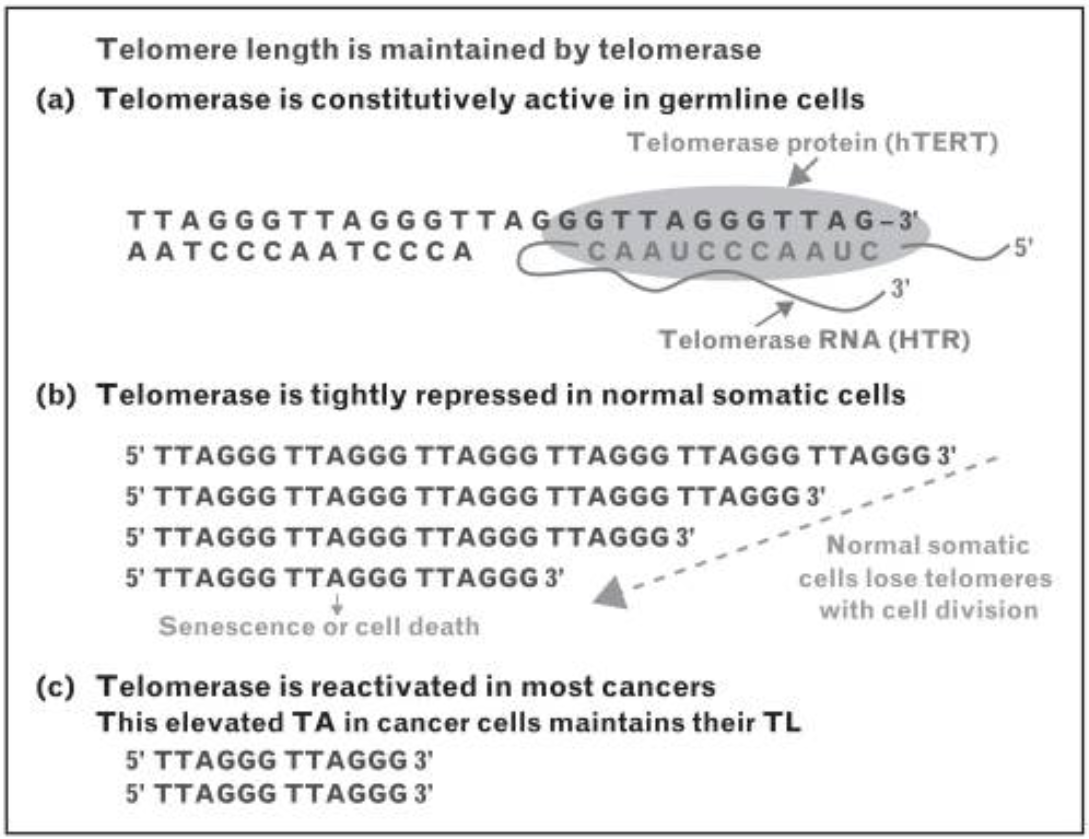
Çok-hücreli ökaryotik örgenliklerin çoğunda telomeraz yalnızca germ hücrelerinde, belli kök hücre tiplerinde (embrionik) ve belli akyuvar tiplerinde etkindir.
Ökaryotik telomerler normal olarak 3′ tek-telli-DNA çıkıntısı ile sonlanır ki, telomer bakımı ve keplenmesi için zorunludur.
Telomerler telomer ilmekleri ya da T-ilmekleri denilen büyük yapılar oluşturur.
T-ilmeklerinde tek-telli DNA uzun bir daire çevresinde kıvrılır ve telomere bağlı proteinleri sağlamlaştırır.
T-ilmeğinin en sonunda tek-telli telomer DNAsı çift-telli bir RNA bölgesi üzerine tutunur ve bunu çift-sarmallı DNAyı bozarak iki telden biri ile baz eşleşmesi kuran telomer teli yoluyla yapar.
Bu üç-telli yapıya bir yer-değiştirme ilmeği ya da D-ilmeği denir (D = displacement).
🛑 TELOMER KISALMASI
TELOMER KISALMASI
Ökaryotlar DNAlarının ucunda yineleyici, kodlama-yapmayan ve telomerler denilen diziler taşır. Telomer kepler bu kromozomlarda açık renk ile gösterildi.
Ökaryotik hücreler doğrusal DNA moleküllerinin uçlarını eşlemlemede özel bir sorun ile karşılaşırlar.
Geciken telin 5' ucundaki RNA primerin kaldırılmasının ardından geriye kalan boşluğu DNA ile doldurmak olanaksızdır.
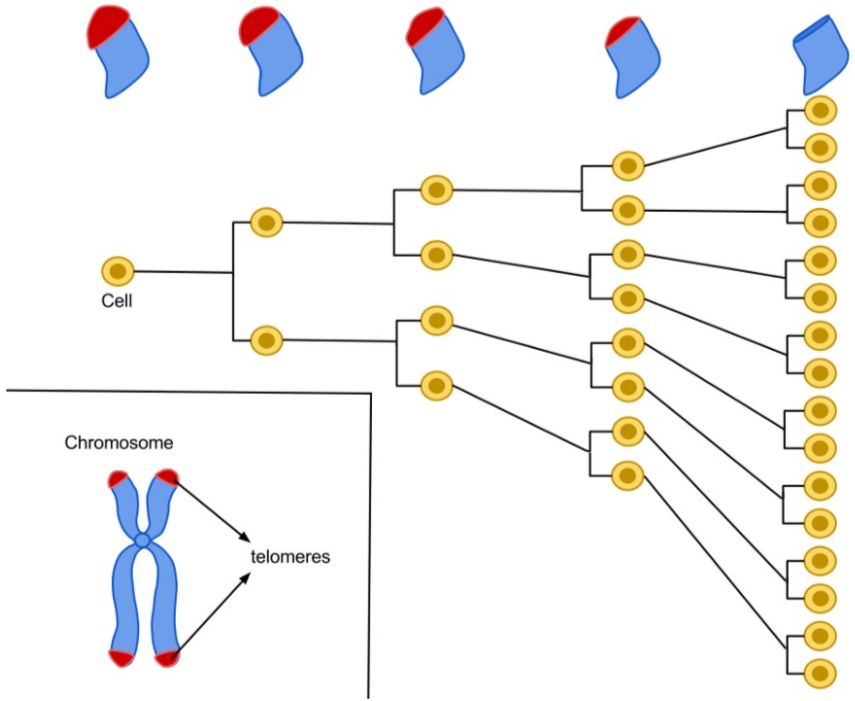
Bu nedenle insanda birçok normal somatik hücrede telomerler her ardışık hücre bölünmesi ile kısalır.
Telomerler kritik bir uzunluğun ötesinde kısalınca, hücre bölünme yapamaz ve yaşlı olarak nitelenir.
Tohum hücrelerinde ve başka kök hücrelerde ve ayrıca kanser hücrelerinde telomerler kısalmaz ve hücreler yaşlanmaz.
Bunun nedeni bu hücrelerde telomer uzunluğunu sürdüren ve telomeraz denilen bir ribonükleoproteinin bulunmasıdır.
Telomerazın normal kısalması somatik hücrelerin uğrayabileceği bölünmelerin sayısını sınırlayarak örgenlikleri kanserden koruyor olabilir.
Büyük tümörlerdeki hücreler uğradıkları bölünmelerin çokluğundan ötürü sık sık olağandışı kısa telomerleri kapsar.
Telomeraz etkinliği kanserli somatik hücrelerde olağandışı yüksektir ve bu kanser hücrelerinin sürmelerini kolaylaştırıyor olabilir.
Birçok kanser hücresi sınırsız hücre bölünmesine yetenekli görünür (“ölümsüz” denilen kültür hücreleri gibi).
Telomeraz etkinliğinin engellenmesinin olanaklı bir kanser sağaltımı olup olmadığı henüz araştırma konusudur.
🛑 SHELTERIN KARMAŞASI
SHELTERIN KARMAŞASI
The Shelterin complex: Characterisation of the proteins that make up the Shelterin complex
Protein Name
Interactions
Function
Telomere repeat binding factor 1 (TERF1 also known as TRF1)
Direct interaction with double stranded TTAGGG repeats
Regulation of telomere length
Telomere repeat binding factor 2 (TERF2 also known as TRF2)
Direct interaction with double stranded TTAGGG repeats
Stabilisation of the T-loop and regulation of telomere length
TERF1 interacting nuclear factor 2 (TINF2 also known as TIN2)
Associates directly with TERF1, TERF2 and ACD and indirectly with POT1
Tethering of ACD and POT1 to TERF1 and TERF2 and tethering TERF1 to TERF2, which stabilises the association of TERF2 with the telomere. Also regulates telomere length
Protection of Telomeres 1 (POT1)
Direct interaction with single stranded telomere overhang
Inhibition of DNA damage response and regulation of telomere length
Shelterin complex subunit and telomerase recruitment factor (ACD, also known as TPP1)
Interaction with TINF2 and POT1
Enhances POT1 binding to single stranded telomere DNA and regulates telomere length in combination with POT1
TERF2 interacting protein (TERF2IP also known as RAP1)
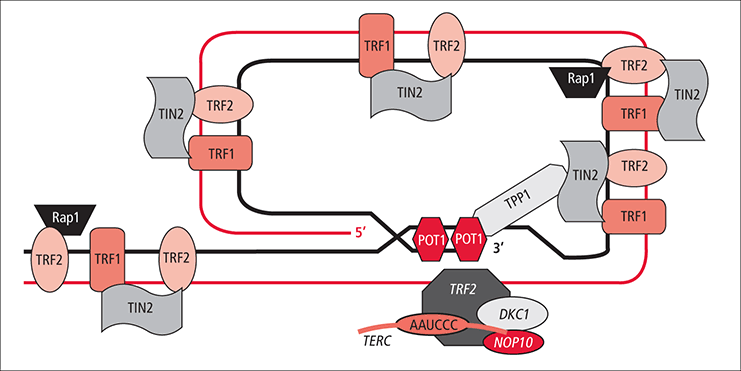
İnsan telomer karmaşası ardışık olarak yineleyen bir DNA dizisinin kromozomal uç zincirinden ve bu zincire bağlanan koruyucu shelterin karmaşasından oluşur. Ek olarak, telomerik DNA yinelemesinin çıkıntı yapan tek-telli uç bölgesi üzerinde koruyucu proteinler bulunur. Bu yalınlaştırılmış şemada protein yapılarının ya da telomerik karmaşanın mimarisinin ayrıntıları göstermedi. (LINK)
Telomer ile birleşik olarak bulunan birçok protein arasında bir bölümü DNA hasar karşılama düzeneklerinde işlevseldir (örneğin: DNA protein kinaz (DNA-PK), p53, poliadenozin difosfat riboz polimeraz (PARP), Tankiraz 1 ve 2, kesilme onarım çapraz-tümleyici ve xeroderma pigmentosum F grubu (ERCC/XPF) radyasyon 51 (RAD51), werner (WRN) ve bloom (BLM)) .
Lamin ilişkili proteinler (LAPlar) ve sessiz bilişim düzenleyici (Sir) proteinleri gibi başka proteinler çekirdek örgütlenmesi ile ilgilidir (ve ayrıca telomer uzunluğunu da denetlerler).
Bu proteinlerin telomer dizisinde bulunuşu ve eylemi büyük ölçüde Shelterin karmaşasını oluşturan proteinler tarafından yönetilir.
Shelterin karmaşası tam işlevsel bir kepleme yapısı oluşturmak üzere telomer yapısı ile birleşmiş altı özelleşmiş proteinden yapılıdır.
SHELTERIN VE TELOMER
Shelterin karmaşasının üyeleri ve telomer DNA dizisi arasındaki ilişkiler telomer yapısını sağlamlaştırır ve RNA onarımı ve uzatımı ile ilgili proteinlerin girişini düzenler.
Bu özelleşmiş nükleoprotein yapı toplu olarak kromozom uçlarında bir tür kapak oluşturma işlevi görür.
Başlıca iki işi yerine getirir: 1) Kromozom kollarının uçlarını uygunsuz DNA onarım düzeneğinden korur (ki bu düzenek gevşek DNA tellerini bir çift sarmal kırılması olarak tanıyabilir ve ortaya kromozom kaynaşmaları çıkabilir); 2) Bu kapaklı yapı tam olmayan DNA eşleniminin bir sonucu olarak kromozom kollarının yakınındaki genlerin bozunumunu önler.
📘 Telomere Biology and Human Phenotype / Kara J. Turner, Vimal Vasu, and Darren K. Griffin (“Cells,” 2019)
📘 Human telomere biology — A contributory and interactive factor in aging, disease risks, and protection / Elizabeth H. Blackburn1, Elissa S. Epel, Jue Lin (“Science, ”2015)
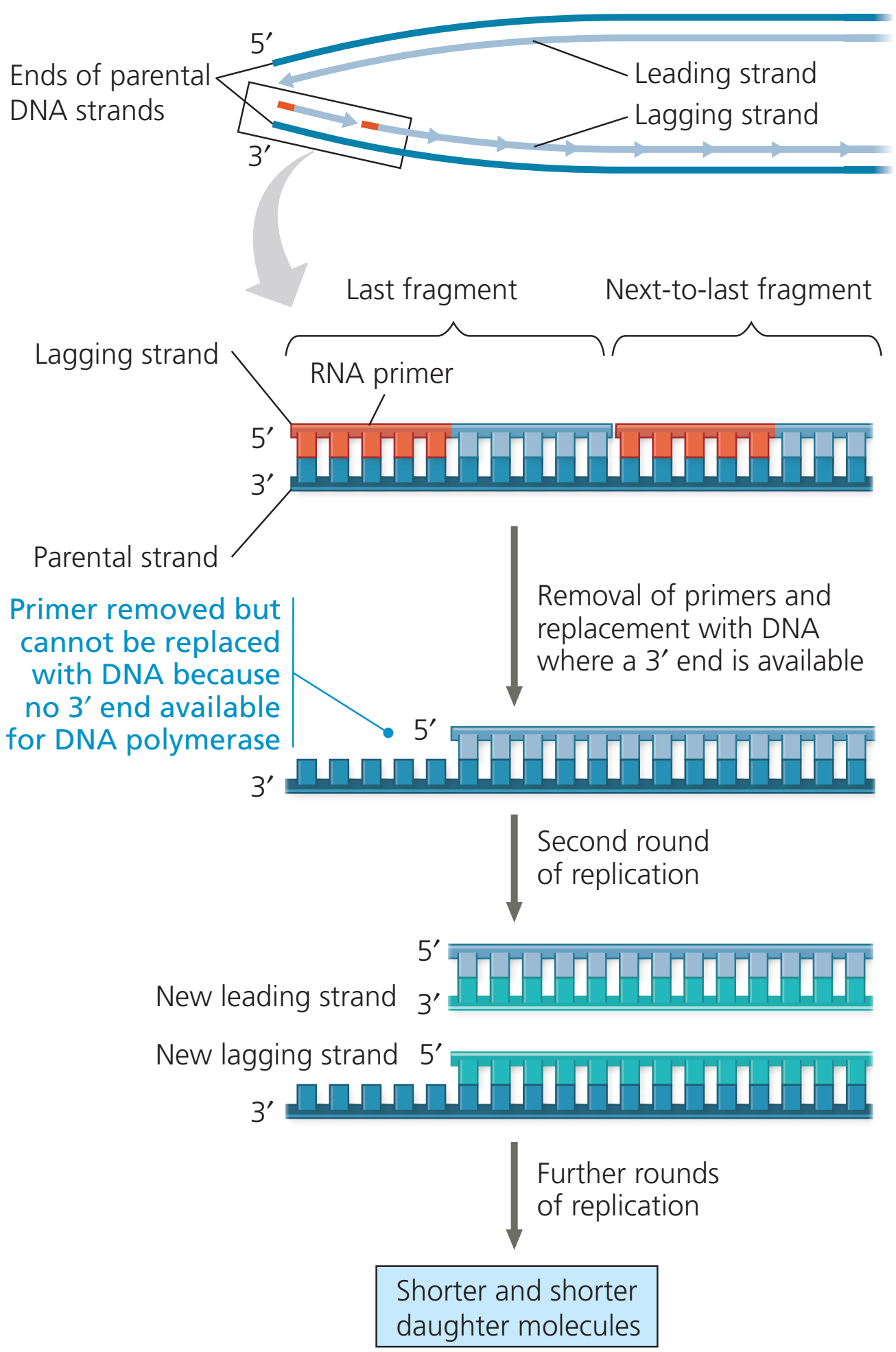
The ends of linear chromosomes pose unique problems during DNA replication. Because DNA polymerases can only elongate from a free 3' hydroxyl group, the replication machinery builds the lagging strand by a backstitching mechanism. RNA primers provide 3'-hydroxyl groups at regular intervals along the lagging strand template. Whereas the leading strand elongates continuously in the 5'-to-3' direction all the way to the end of the template, the lagging strand stops short of the end. Even if a final RNA primer were built at the very end of the chromosome, the lagging strand would not be complete. The final primer would provide a 3'-OH group to synthesize DNA, but the primers would later need to be removed. The 3'-hydroxyl groups on adjacent DNA fragments provide starting places for replacing the RNA with DNA. However, at the end of the chromosome there is no 3'-OH group available to prime DNA synthesis. Because of this inability to replicate the ends, chromosomes would progressively shorten during each replication cycle. This "end-replication" problem is solved by the enzyme telomerase. The ends of chromosomes contain a G-rich series of repeats called a telomere. Telomerase recognizes the tip of an existing repeat sequence. Using an RNA template within the enzyme, telomerase elongates the parental strand in the 5'-to-3' direction, and adds additional repeats as it moves down the parental strand. The lagging strand is then completed by DNA polymerase alpha, which carries a DNA primase as one of its subunits. In this way, the original information at the ends of linear chromosomes is completely copied in the new DNA.
📹 Telomeres and cell senescence / Khan Academy (VİDEO)
📹 Telomeres and cell senescence / Khan Academy (LINK)
📹 A full explanation about the Telomerase and the end replication problem / Biomedical and Biological Sciences (VİDEO)
📹 A full explanation about the Telomerase and the end replication problem / Biomedical and Biological Sciences (LINK)
This video explains ehats is the telomeres, the telomerase activity, and the end replication problem,
Elizabeth Blackburn (UCSF) | Discovery of Telomeric DNA and Telomerase
📹 Elizabeth Blackburn (UCSF) | Discovery of Telomeric DNA and Telomerase
Human chromosomes (grey) capped by telomeres (white).
A telomere is a region of repetitive nucleotide sequences at each end of a chromosome, which protects the end of the chromosome from deterioration or from fusion with neighboring chromosomes. Its name is derived from the Greek nouns telos (τέλος) "end" and merοs (μέρος, root: μερ-) "part". For vertebrates, the sequence of nucleotides in telomeres is 5′-TTAGGG-3′, with the complementary DNA strand being 3′-AATCCC-5′, with a single-stranded TTAGGG overhang. This sequence of TTAGGG is repeated approximately 2,500 times in humans. In humans, average telomere length declines from about 11 kilobases at birth to fewer than 4 kilobases in old age, with the average rate of decline being greater in men than in women.
During chromosome replication, the enzymes that duplicate DNA cannot continue their duplication all the way to the end of a chromosome, so in each duplication the end of the chromosome is shortened (this is because the synthesis of Okazaki fragments requires RNA primers attaching ahead on the lagging strand). The telomeres are disposable buffers at the ends of chromosomes which are truncated during cell division; their presence protects the genes before them on the chromosome from being truncated instead. The telomeres themselves are protected by a complex of shelterin proteins, as well as by the RNA that telomeric DNA encodes (TERRA).
Over time, due to each cell division, the telomere ends become shorter. They are replenished by an enzyme, telomerase reverse transcriptase.
In the early 1970s, Russian theorist Alexei Olovnikov first recognized that chromosomes could not completely replicate their ends. Building on this, and to accommodate Leonard Hayflick's idea of limited somatic cell division, Olovnikov suggested that DNA sequences are lost every time a cell replicates until the loss reaches a critical level, at which point cell division ends.
In 1983, Barbara McClintock, an American cytogeneticist and the first woman to receive an unshared Nobel Prize in Physiology or Medicine, received the Nobel Prize for observing that the chromosomes lacking end parts became “sticky” and hypothesized the existence of a special structure at the chromosome tip that would maintain chromosome stability.
Telomeres are found at the termini of chromosomes. The end of a telomere inserts back into the main body of the telomere to form a T-loop
Telomeres are repetitive nucleotide sequences located at the termini of linear chromosomes of most eukaryotic organisms. For vertebrates, the sequence of nucleotides in telomeres is TTAGGG. Most prokaryotes, having circular chromosomes rather than linear, do not have telomeres. Telomeres compensate for incomplete semi-conservative DNA replication at chromosomal ends. A protein complex known as shelterin serves to protect the ends of telomeres from being recognised as double-strand breaks by inhibiting homologous recombination (HR) and non-homologous end joining (NHEJ).
In most prokaryotes, chromosomes are circular and thus do not have ends to suffer premature replication termination. A small fraction of bacterial chromosomes (such as those in Streptomyces, Agrobacterium, and Borrelia) are linear and possess telomeres, which are very different from those of the eukaryotic chromosomes in structure and functions. The known structures of bacterial telomeres take the form of proteins bound to the ends of linear chromosomes, or hairpin loops of single-stranded DNA at the ends of the linear chromosomes.
While replicating DNA, the eukaryotic DNA replication enzymes (the DNA polymerase protein complex) cannot replicate the sequences present at the ends of the chromosomes (or more precisely the chromatid fibres). Hence, these sequences and the information they carry may get lost. This is the reason telomeres are so important in context of successful cell division: They "cap" the end-sequences and themselves get lost in the process of DNA replication. But the cell has an enzyme called telomerase, which carries out the task of adding repetitive nucleotide sequences to the ends of the DNA. Telomerase "replenishes" the telomere "cap." In most multicellular eukaryotic organisms, telomerase is active only in germ cells, some types of stem cells such as embryonic stem cells, and certain white blood cells. Telomerase can be reactivated and telomeres reset back to an embryonic state by somatic cell nuclear transfer. The steady shortening of telomeres with each replication in somatic (body) cells may have a role in senescence and in the prevention of cancer. This is because the telomeres act as a sort of time-delay "fuse", eventually running out after a certain number of cell divisions and resulting in the eventual loss of vital genetic information from the cell's chromosome with future divisions.
Telomere length varies greatly between species, from approximately 300 base pairs in yeast to many kilobases in humans, and usually is composed of arrays of guanine-rich, six- to eight-base-pair-long repeats. Eukaryotic telomeres normally terminate with 3′ single-stranded-DNA overhang, which is essential for telomere maintenance and capping. Multiple proteins binding single- and double-stranded telomere DNA have been identified. These function in both telomere maintenance and capping. Telomeres form large loop structures called telomere loops, or T-loops. Here, the single-stranded DNA curls around in a long circle, stabilized by telomere-binding proteins. At the very end of the T-loop, the single-stranded telomere DNA is held onto a region of double-stranded DNA by the telomere strand disrupting the double-helical DNA, and base pairing to one of the two strands. This triple-stranded structure is called a displacement loop or D-loop.
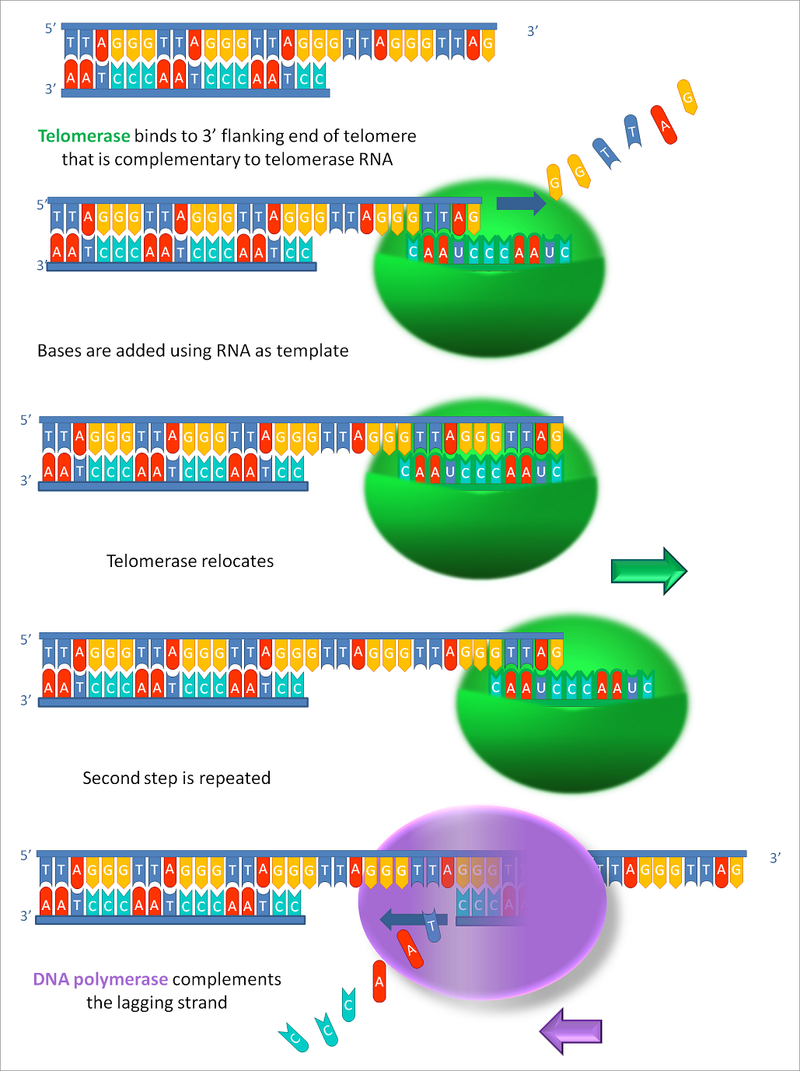
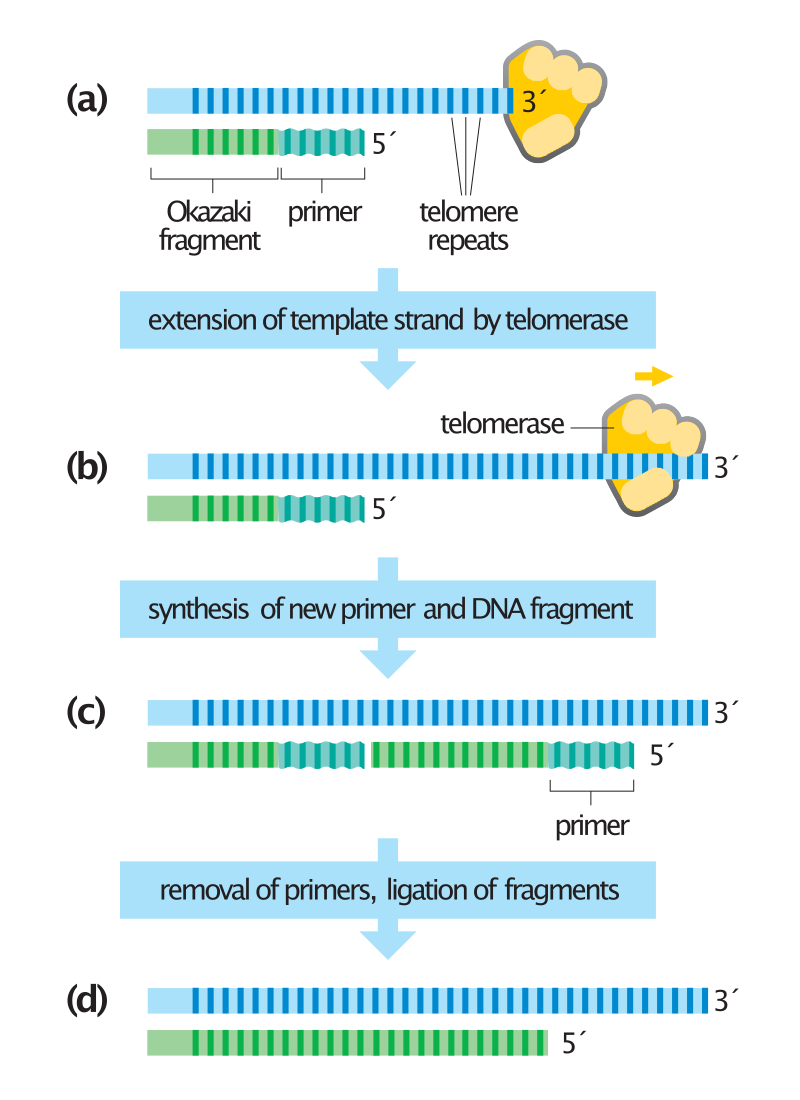
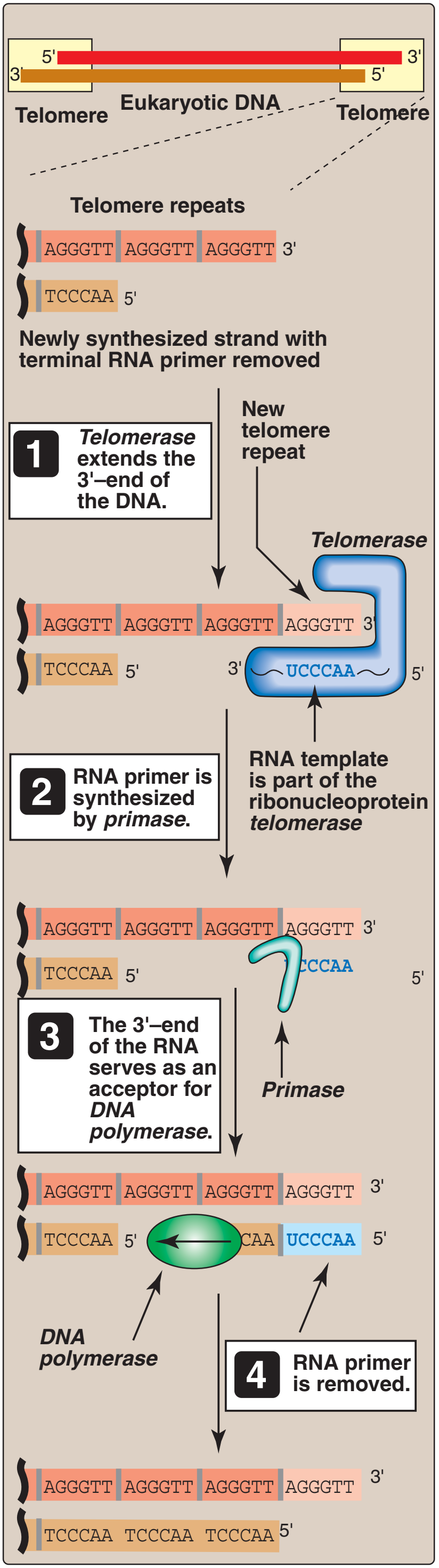
Synthesis of chromosome ends by telomerase.
Telomeres at the ends of chromosomes contain many repeats of a short DNA sequence. (a) When the last Okazaki fragment on the lagging strand is completed, a short region of DNA remains unduplicated at the chromosome tip. (b) To avoid chromosome shortening, the enzyme telomerase extends the template strand by synthesizing additional copies of the telomere repeat sequence. Telomerase does not need a DNA template to perform this reaction because it contains its own template (made of RNA). (c) The extended DNA template is used by the DNA polymerase machinery to synthesize a new Okazaki fragment. (d) Primers are removed, and the internal primer is replaced with DNA as usual. The last primer cannot be replaced with DNA because not enough template strand is present. A short stretch of single-stranded DNA therefore remains at the end of the chromosome. In this example, the length of the chromosome has increased slightly during DNA synthesis; normally, telomeres are maintained at a constant length by complex regulatory mechanisms. In some cells, telomerase is absent and telomeres gradually shorten with each cell division.
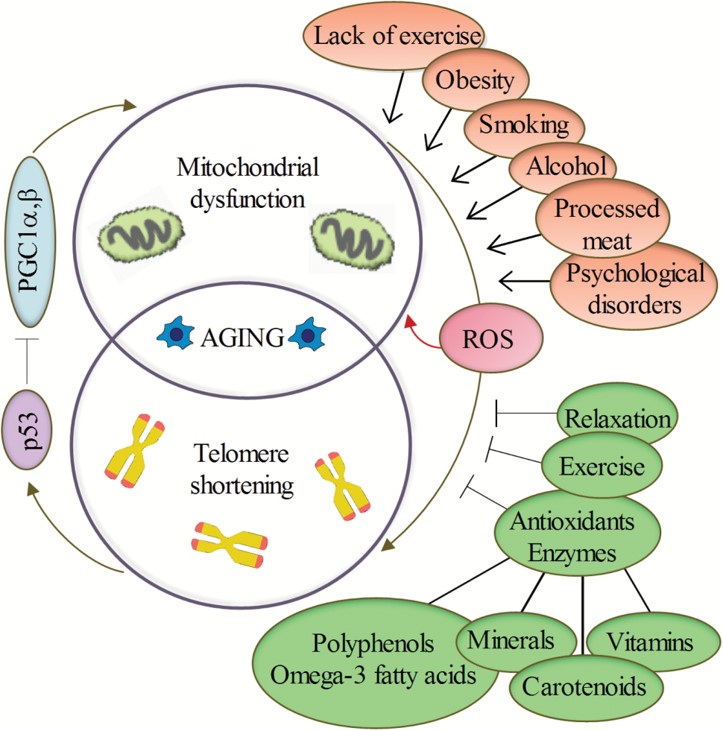
Telomere shortening in humans can induce replicative senescence, which blocks cell division. This mechanism appears to prevent genomic instability and development of cancer in human aged cells by limiting the number of cell divisions. However, shortened telomeres impair immune function that might also increase cancer susceptibility. If telomeres become too short, they have the potential to unfold from their presumed closed structure. The cell may detect this uncapping as DNA damage and then either stop growing, enter cellular old age (senescence), or begin programmed cell self-destruction (apoptosis) depending on the cell's genetic background (p53 status). Uncapped telomeres also result in chromosomal fusions. Since this damage cannot be repaired in normal somatic cells, the cell may even go into apoptosis. Many aging-related diseases are linked to shortened telomeres. Organs deteriorate as more and more of their cells die off or enter cellular senescence.
At the very distal end of the telomere is a 300 base pair single-stranded portion, which forms the T-loop. This loop is analogous to a knot, which stabilizes the telomere, preventing the telomere ends from being recognized as break points by the DNA repair machinery. Should non-homologous end joining occur at the telomeric ends, chromosomal fusion will result. The T-loop is held together by several proteins, the most notable ones being TRF1,TRF2,POT1, RAP1, and TIN2, collectively referred to as the shelterin complex. In humans, the shelterin complex consists of six proteins identified as TRF1,TRF2,TIN2,POT1,TPP1, and RAP1.
Shelterin co-ordinates the T-loop formation of telomeres. Schematics of the arrangement of proteins in the telosome/shelterin complex.
Telomeres shorten in part because of the end replication problem that is exhibited during DNA replication in eukaryotes only. Because DNA replication does not begin at either end of the DNA strand, but starts in the center, and considering that all known DNA polymerases read the template strand in the 3' to 5' direction, one finds a leading and a lagging strand on the DNA molecule being replicated.
On the leading strand, DNA polymerase can make a complementary DNA strand without any difficulty because it reads the template strand from 3' to 5'. However, there is a problem going in the other direction on the lagging strand. To counter this, short sequences of RNA acting as primers attach to the lagging strand a short distance ahead of where the initiation site was. The DNA polymerase can start replication at that point and go to the end of the initiation site. This causes the formation of Okazaki fragments. More RNA primers attach further on the DNA strand and DNA polymerase comes along and continues to make a new DNA strand.
Lagging strand during DNA replication.
Eventually, the last RNA primer attaches, and DNA polymerase, RNA nuclease, and DNA ligase come along to convert the RNA (of the primers) to DNA and to seal the gaps in between the Okazaki fragments. But, in order to change RNA to DNA, there must be another DNA strand in front of the RNA primer. This happens at all the sites of the lagging strand, but it does not happen at the end where the last RNA primer is attached. Ultimately, that RNA is destroyed by enzymes that degrade any RNA left on the DNA. Thus, a section of the telomere is lost during each cycle of replication at the 5' end of the lagging strand's daughter.
However, test-tube studies have shown that telomeres are highly susceptible to oxidative stress. There is evidence that oxidative stress-mediated DNA damage is an important determinant of telomere shortening. Telomere shortening due to free radicals explains the difference between the estimated loss per division because of the end-replication problem (c. 20 bp) and actual telomere shortening rates (50–100 bp), and has a greater absolute impact on telomere length than shortening caused by the end-replication problem. Population-based studies have also indicated an interaction between anti-oxidant intake and telomere length. In the Long Island Breast Cancer Study Project (LIBCSP), authors found a moderate increase in breast cancer risk among women with the shortest telomeres and lower dietary intake of beta carotene, vitamin C or E. These results suggest that cancer risk due to telomere shortening may interact with other mechanisms of DNA damage, specifically oxidative stress.
Telomere shortening is associated with aging, mortality and aging-related diseases. Normal aging is associated with telomere shortening in both humans and mice, and studies on genetically modified animal models suggest causal links between telomere erosion and aging. However, it is not known whether short telomeres are just a sign of cellular age or if they themselves actually contribute to the aging process.
The age of a father plays a role in the length of a child’s telomeres, which has evolutionary implications. Although leukocyte telomeres shorten with age, sperm telomeres lengthen with age. Shorter telomeres are theorized to impose lower energy costs (due to less replication) but also have immune system-related and other aging- and disease-related costs, so the effect of paternal age on telomere length might be an adaptation to increase the chances that the child will be fit for the environment they’re born into.
Potential effect of psychological stress
Potential effect of psychological stress (W) Meta-analyses found that increased perceived psychological stress was associated with a small decrease in telomere length—but that these associations attenuate to no significant association when accounting for publication bias. The literature concerning telomeres as integrative biomarkers of exposure to stress and adversity is dominated by cross-sectional and correlational studies, which makes causal interpretation problematic. A 2020 review argued that the relationship between psychosocial stress and telomere length appears strongest for stress experienced in utero or early life.
The phenomenon of limited cellular division was first observed by Leonard Hayflick, and is now referred to as the Hayflick limit. Significant discoveries were subsequently made by a group of scientists organized at Geron Corporation by Geron's founder Michael D. West, that tied telomere shortening with the Hayflick limit. The cloning of the catalytic component of telomerase enabled experiments to test whether the expression of telomerase at levels sufficient to prevent telomere shortening was capable of immortalizing human cells. Telomerase was demonstrated in a 1998 publication in Science to be capable of extending cell lifespan, and now is well-recognized as capable of immortalizing human somatic cells.
The average cell will divide between 50 and 70 times before cell death. As the cell divides the telomeres on the end of the chromosome get smaller. The Hayflick limit is the theoretical limit to the number of times a cell may divide until the telomere becomes so short that division is inhibited and the cell enters senescence.
The average cell will divide between 50-70 times before cell death. As the cell divides the telomeres on the end of the chromosome get smaller. The Hayflick Limit is the theory that due to the telomeres shortening through each division, the telomeres will eventually no longer be present on the chromosome. This end stage is known as senescence and proves the concept that links the deterioration of telomeres and aging.
It is becoming apparent that reversing shortening of telomeres through temporary activation of telomerase may be a potent means to slow aging. The reason that this would extend human life is because it would extend the Hayflick limit. Three routes have been proposed to reverse telomere shortening: drugs, gene therapy, or metabolic suppression, so-called torpor/hibernation. So far these ideas have not been proven in humans, but it has been demonstrated that telomere shortening is reversed in hibernation and aging is slowed (Turbill, et al. 2012 & 2013) and that hibernation prolongs life-span (Lyman et al. 1981). It has also been demonstrated that telomere extension has successfully reversed some signs of aging in laboratory mice and the nematode worm species Caenorhabditis elegans. It has been hypothesized that longer telomeres and especially telomerase activation might cause increased cancer (e.g. Weinstein and Ciszek, 2002). However, longer telomeres might also protect against cancer, because short telomeres are associated with cancer. It has also been suggested that longer telomeres might cause increased energy consumption.
Techniques to extend telomeres could be useful for tissue engineering, because they might permit healthy, noncancerous mammalian cells to be cultured in amounts large enough to be engineering materials for biomedical repairs.
Two studies on long-lived seabirds demonstrate that the role of telomeres is far from being understood. In 2003, scientists observed that the telomeres of Leach's storm-petrel (Oceanodroma leucorhoa) seem to lengthen with chronological age, the first observed instance of such behaviour of telomeres. In 2006, Juola et al. reported that in another unrelated, long-lived seabird species, the great frigatebird (Fregata minor), telomere length did decrease until at least c. 40 years of age (i.e. probably over the entire lifespan), but the speed of decrease slowed down massively with increasing ages, and that rates of telomere length decrease varied strongly between individual birds. They concluded that in this species (and probably in frigatebirds and their relatives in general), telomere length could not be used to determine a bird's age sufficiently well. Thus, it seems that there is much more variation in the behavior of telomere length than initially believed.
Furthermore, Gomes et al. found, in a study of the comparative biology of mammalian telomeres, that telomere length of different mammalian species correlates inversely, rather than directly, with lifespan, and they concluded that the contribution of telomere length to lifespan remains controversial. Harris et al. found little evidence that, in humans, telomere length is a significant biomarker of normal aging with respect to important cognitive and physical abilities. Gilley and Blackburn tested whether cellular senescence in paramecium is caused by telomere shortening, and found that telomeres were not shortened during senescence.
Telomeres are critical for maintaining genomic integrity and may be factors for age-related diseases. Laboratory studies show that telomere dysfunction or shortening is commonly acquired due process of cellular aging and tumor development. Short telomeres can lead to genomic instability, chromosome loss and the formation of non-reciprocal translocations; and telomeres in tumor cells and their precursor lesions are significantly shorter than surrounding normal tissue.
Observational studies have found shortened telomeres in many types of experimental cancers. In addition, people with cancer have been found to possess shorter leukocyte telomeres than healthy controls. Recent meta-analyses suggest 1.4 to 3.0 fold increased risk of cancer for those with the shortest vs. longest telomeres. However, the increase in risk varies by age, sex, tumor type, and differences in lifestyle factors.
Several techniques are currently employed to assess average telomere length in eukaryotic cells. One method is the Terminal Restriction Fragment (TRF) southern blot. A Real-Time PCR assay for telomere length involves determining the Telomere-to-Single Copy Gene (T/S)ratio, which is demonstrated to be proportional to the average telomere length in a cell.
Tools have also been developed to estimate the length of telomere from whole genome sequencing (WGS) experiments. Amongst these are TelSeq, telomerecat and telomereHunter. Length estimation from WGS typically works by differentiating telomere sequencing reads and then inferring the length of telomere that produced that number of reads. These methods have been shown to correlate with preexisting methods of estimation such as PCR and TRF. Flow-FISH is used to quantify the length of telomeres in human white blood cells. A semi-automated method for measuring the average length of telomeres with Flow FISH was published in Nature Protocols in 2006.
While multiple companies offer telomere length measurement services, the utility of these measurements for widespread clinical or personal use has been questioned. Nobel Prize winner Elizabeth Blackburn, who was co-founder of one company, promoted the clinical utility of telomere length measures.
Ectothermic telomeres
Ectothermic telomeres
Ectothermic telomeres (W)
Most research on telomere length and regulation, and its relationship to cancer and aging, has been performed on mammals, especially humans, which have little or no somatic telomerase production. Ectotherms are significantly more likely than endotherms to have variation in somatic telomerase expression. For instance, in many fish, telomerase occurs throughout the body (and associated with this, telomere length is roughly the same across all its tissue). Studies on ectotherms, and other non-mammalian organisms, show that there is no single universal model of telomere erosion; rather, there is wide variation in relevant dynamics across Metazoa, and even within smaller taxonomic groups these patterns appear diverse. Due to the different reproductive timelines of some ectotherms, selection on disease is relevant for a much larger fraction of these creatures’ lives than it is for mammals, so early- and late-life telomere length, and their possible links to cancer, seem especially important in these species from a life history theory point of view.
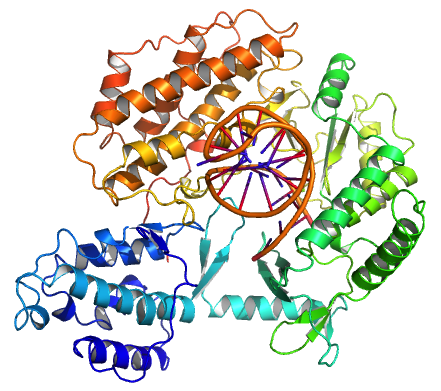
Tribolium castaneum telomerase catalytic subunit, TERT, bound to putative RNA template and telomeric DNA (PDB 3KYL)
Tribolium castaneum telomerase catalytic subunit, TERT, bound to an RNA-DNA hairpin designed to resemble the putative RNA-templating region and telomeric DNA.
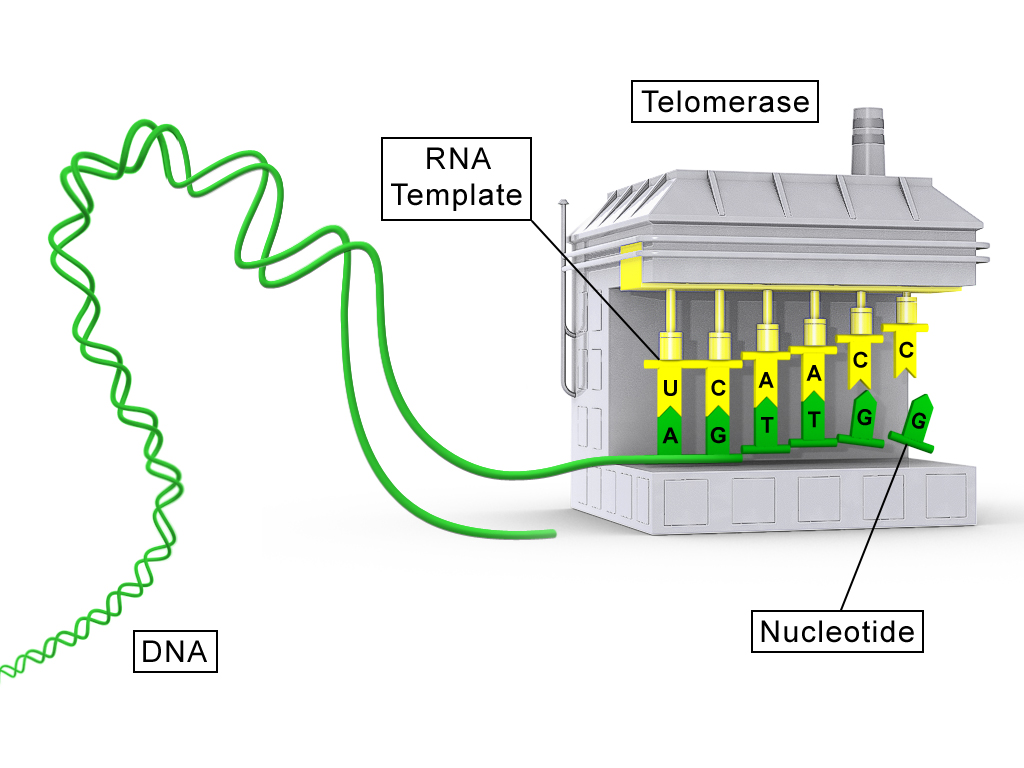
An illustration of a telomerase molecule.
A conceptual diagram showing the protein component of telomerase (TERT) in grey and the RNA component (TR) in yellow .
The existence of a compensatory mechanism for telomere shortening was first found by Soviet biologist Alexey Olovnikov in 1973, who also suggested the telomere hypothesis of aging and the telomere's connections to cancer.
The role of telomeres and telomerase in cell aging and cancer was established by scientists at biotechnology company Geron with the cloning of the RNA and catalytic components of human telomerase and the development of a polymerase chain reaction (PCR) based assay for telomerase activity called the TRAP assay, which surveys telomerase activity in multiple types of cancer.
The negative stain electron microscopy (EM) structures of human and Tetrahymena telomerases were characterized in 2013. Two years later, the first cryo-electron microscopy (cryo-EM) structure of telomerase holoenzyme (Tetrahymena) was determined. In 2018, the structure of human telomerase was determined through cryo-EM by UC Berkeley scientists.
The molecular composition of the human telomerase complex was determined by Scott Cohen and his team at the Children's Medical Research Institute (Sydney Australia) and consists of two molecules each of human telomerase reverse transcriptase (TERT), telomerase RNA (TR or TERC), and dyskerin (DKC1). The genes of telomerase subunits, which include TERT, TERC, DKC1 and TEP1, are located on different chromosomes. The human TERT gene (hTERT) is translated into a protein of 1132 amino acids. TERT polypeptide folds with (and carries) TERC, a non-coding RNA (451 nucleotides long). TERT has a 'mitten' structure that allows it to wrap around the chromosome to add single-stranded telomere repeats.
TERT is a reverse transcriptase, which is a class of enzyme that creates single-stranded DNA using single-stranded RNA as a template.
The protein consists of four conserved domains (RNA-Binding Domain (TRBD), fingers, palm and thumb), organized into a ring configuration that shares common features with retroviral reverse transcriptases, viral RNA polymerases and bacteriophage B-family DNA polymerases.
TERT proteins from many eukaryotes have been sequenced.
By using TERC, TERT can add a six-nucleotide repeating sequence, 5'-TTAGGG (in vertebrates, the sequence differs in other organisms) to the 3' strand of chromosomes. These TTAGGG repeats (with their various protein binding partners) are called telomeres. The template region of TERC is 3'-CAAUCCCAAUC-5'.
Telomerase can bind the first few nucleotides of the template to the last telomere sequence on the chromosome, add a new telomere repeat (5'-GGTTAG-3') sequence, let go, realign the new 3'-end of telomere to the template, and repeat the process. Telomerase reverses telomere shortening.
An image illustrating how telomerase elongates telomere ends progressively.
Telomerase restores short bits of DNA known as telomeres, which are otherwise shortened when a cell divides via mitosis.
In normal circumstances, where telomerase is absent, if a cell divides recursively, at some point the progeny reach their Hayflick limit, which is believed to be between 50–70 cell divisions. At the limit the cells become senescent and cell division stops. Telomerase allows each offspring to replace the lost bit of DNA, allowing the cell line to divide without ever reaching the limit. This same unbounded growth is a feature of cancerous growth.
Embryonic stem cells express telomerase, which allows them to divide repeatedly and form the individual. In adults, telomerase is highly expressed only in cells that need to divide regularly, especially in male sperm cells but also in epidermal cells, in activated T cell and B celllymphocytes, as well as in certain adult stem cells, but in the great majority of cases somatic cells do not express telomerase.
A comparative biology study of mammalian telomeres indicated that telomere length of some mammalian species correlates inversely, rather than directly, with lifespan, and concluded that the contribution of telomere length to lifespan is unresolved. Telomere shortening does not occur with age in some postmitotic tissues, such as in the rat brain. In humans, skeletal muscle telomere lengths remain stable from ages 23 –74. In baboon skeletal muscle, which consists of fully differentiated post-mitotic cells, less than 3% of myonuclei contain damaged telomeres and this percentage does not increase with age. Thus, telomere shortening does not appear to be a major factor in the aging of the differentiated cells of brain or skeletal muscle. In human liver, cholangiocytes and hepatocytes show no age-related telomere shortening. Another study found little evidence that, in humans, telomere length is a significant biomarker of normal aging with respect to important cognitive and physical abilities.
Some experiments have raised questions on whether telomerase can be used as an anti-aging therapy, namely, the fact that mice with elevated levels of telomerase have higher cancer incidence and hence do not live longer. On the other hand, one study showed that activating telomerase in cancer-resistant mice by overexpressing its catalytic subunit extended lifespan.
A study that focused on Ashkenazi Jews found that long-lived subjects inherited a hyperactive version of telomerase.
In vitro, when cells approach the Hayflick limit, the time to senescence can be extended by inactivating the tumor suppressor proteins - p53 and Retinoblastoma protein (pRb). Cells that have been so-altered eventually undergo an event termed a "crisis" when the majority of the cells in the culture die. Sometimes, a cell does not stop dividing once it reaches a crisis. In a typical situation, the telomeres are shortened and chromosomal integrity declines with every subsequent cell division. Exposed chromosome ends are interpreted as double-stranded breaks (DSB) in DNA; such damage is usually repaired by reattaching (relegating) the broken ends together. When the cell does this due to telomere-shortening, the ends of different chromosomes can be attached to each other. This solves the problem of lacking telomeres, but during cell division anaphase, the fused chromosomes are randomly ripped apart, causing many mutations and chromosomal abnormalities. As this process continues, the cell's genome becomes unstable. Eventually, either fatal damage is done to the cell's chromosomes (killing it via apoptosis), or an additional mutation that activates telomerase occurs.
With telomerase activation some types of cells and their offspring become immortal (bypass the Hayflick limit), thus avoiding cell death as long as the conditions for their duplication are met. Many cancer cells are considered 'immortal' because telomerase activity allows them to live much longer than any other somatic cell, which, combined with uncontrollable cell proliferation is why they can form tumors. A good example of immortal cancer cells is HeLa cells, which have been used in laboratories as a model cell line since 1951.
While this method of modelling human cancer in cell culture is effective and has been used for many years by scientists, it is also very imprecise. The exact changes that allow for the formation of the tumorigenic clones in the above-described experiment are not clear. Scientists addressed this question by the serial introduction of multiple mutations present in a variety of human cancers. This has led to the identification of mutation combinations that form tumorigenic cells in a variety of cell types. While the combination varies by cell type, the following alterations are required in all cases: TERT activation, loss of p53 pathway function, loss of pRb pathway function, activation of the Ras or mycproto-oncogenes, and aberration of the PP2A protein phosphatase. That is to say, the cell has an activated telomerase, eliminating the process of death by chromosome instability or loss, absence of apoptosis-induction pathways, and continued mitosis activation.
This model of cancer in cell culture accurately describes the role of telomerase in actual human tumors. Telomerase activation has been observed in ~90% of all human tumors, suggesting that the immortality conferred by telomerase plays a key role in cancer development. Of the tumors without TERT activation, most employ a separate pathway to maintain telomere length termed Alternative Lengthening of Telomeres (ALT). The exact mechanism behind telomere maintenance in the ALT pathway is unclear, but likely involves multiple recombination events at the telomere.
Elizabeth Blackburnet al., identified the upregulation of 70 genes known or suspected in cancer growth and spread through the body, and the activation of glycolysis, which enables cancer cells to rapidly use sugar to facilitate their programmed growth rate (roughly the growth rate of a fetus).
Approaches to controlling telomerase and telomeres for cancer therapy include gene therapy,immunotherapy, small-molecule and signal pathway inhibitors.
The ability to maintain functional telomeres may be one mechanism that allows cancer cells to grow in vitro for decades. Telomerase activity is necessary to preserve many cancer types and is inactive in somatic cells, creating the possibility that telomerase inhibition could selectively repress cancer cell growth with minimal side effects. If a drug can inhibit telomerase in cancer cells, the telomeres of successive generations will progressively shorten, limiting tumor growth.
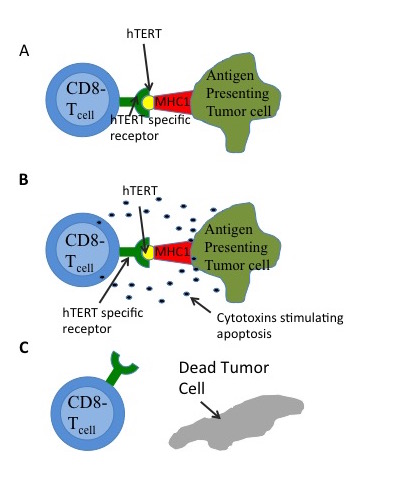
Figure 4:A) Tumor cells expressing hTERT will actively degrade some of the protein and process for presenting. The major histocompatibility complex 1(MHC1), can then present the hTERT epitote. CD8- T cells that have antibodies against hTERT will then bind to the presented epitote. B) As a result of the antigenic binding, the T cells will release cytotoxins, which can be absorbed by the affected cell. C) These cytotoxins induce multiple proteases and results in apoptosis (or cell death).
Telomerase is a good biomarker for cancer detection because most human cancers cells express high levels of it. Telomerase activity can be identified by its catalytic protein domain (hTERT). This is the rate-limiting step in telomerase activity. It is associated with many cancer types. Various cancer cells and fibroblasts transformed with hTERT cDNA have high telomerase activity, while somatic cells do not. Cells testing positive for hTERT have positive nuclear signals. Epithelial stem cell tissue and its early daughter cells are the only noncancerous cells in which hTERT can be detected. Since hTERT expression is dependent only on the number of tumor cells within a sample, the amount of hTERT indicates the severity of a cancer.
The expression of hTERT can also be used to distinguish benign tumors from malignant tumors. Malignant tumors have higher hTERT expression than benign tumors. Real-time reverse transcription polymerase chain reaction (RT-PCR) quantifying hTERT expression in various tumor samples verified this varying expression.
The lack of telomerase does not affect cell growth, until the telomeres are short enough to cause cells to “die or undergo growth arrest”. However, inhibiting telomerase alone is not enough to destroy large tumors. It must be combined with surgery, radiation,chemotherapy or immunotherapy.
Cells may reduce their telomere length by only 50-252 base pairs per cell division, which can lead to a long lag phase.
Immunotherapy
Immunotherapy (W) Immunotherapy successfully treats some kinds of cancer, such as melanoma. This treatment involves manipulating a human's immune system to destroy cancerous cells. Humans have two major antigen identifying lymphocytes:CD8+ cytotoxic T-lymphocytes (CTL) and CD4+ helper T-lymphocytes that can destroy cells. Antigen receptors on CTL can bind to a 9-10 amino acid chain that is presented by the major histocompatibility complex (MHC) as in Figure 4. HTERT is a potential target antigen. Immunotargeting should result in relatively few side effects since hTERT expression is associated only with telomerase and is not essential in almost all somatic cells. GV1001 uses this pathway. Experimental drug and vaccine therapies targeting active telomerase have been tested in mouse models, and clinical trials have begun. One drug, imetelstat, is being clinically researched as a means of interfering with telomerase in cancer cells. Most of the harmful cancer-related effects of telomerase are dependent on an intact RNA template. Cancer stem cells that use an alternative method of telomere maintenance are still killed when telomerase's RNA template is blocked or damaged.
Telomerase Vaccines
Telomerase Vaccines (W)
Two telomerase vaccines have been developed: GRNVAC1 and GV1001. GRNVAC1 isolates dendritic cells and the RNA that codes for the telomerase protein and puts them back into the patient to make cytotoxic T cells that kill the telomerase-active cells. GV1001 is a peptide from the active site of hTERT and is recognized by the immune system that reacts by killing the telomerase-active cells.
Targeted apoptosis
Targeted apoptosis (W)
Another independent approach is to use oligoadenylated anti-telomerase antisense oligonucleotides and ribozymes to target telomerase RNA, reducing dissociation and apoptosis (Figure 5). The fast induction of apoptosis through antisense binding may be a good alternative to the slower telomere shortening.
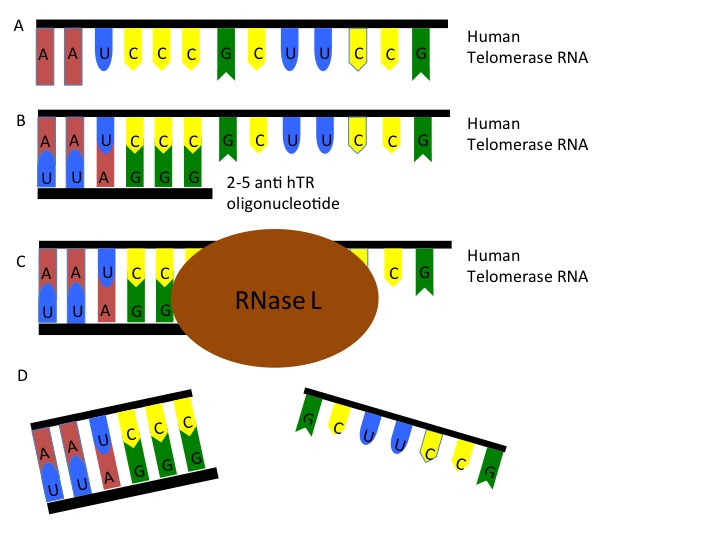
Figure 5: A) Human telomerase RNA (hTR) is present in the cell and can be targeted. B) 2-5 anti-hTR oligonucleotides is a specialized antisense oligo that can bind to the telomerase RNA. C) Once bound, the 2-5 anti-hTR oligonucleotide recruits RNase L to the sequence. Once recruited, the RNase L creates a single cleavage in the RNA (D) and causes dissociation of the RNA sequence.
Small interfering RNA (siRNA)
Small interfering RNA (siRNA) (W) siRNAs are small RNA molecules that induce the sequence-specific degradation of other RNAs. siRNA treatment can function similar to traditional gene therapy by destroying the mRNA products of particular genes, and therefore preventing the expression of those genes. A 2012 study found that targeting TERC with an siRNA reduced telomerase activity by more than 50% and resulted in decreased viability of immortal cancer cells. Treatment with both the siRNA and radiation caused a greater reduction in tumor size in mice than treatment with radiation alone, suggesting that targeting telomerase could be a way to increase the efficacy of radiation in treating radiation-resistant tumors.
Blackburn also discovered that mothers caring for very sick children have shorter telomeres when they report that their emotional stress is at a maximum and that telomerase was active at the site of blockages in coronary artery tissue, possibly accelerating heart attacks.
In 2009, it was shown that the amount of telomerase activity significantly increased following psychological stress. Across the sample of patients telomerase activity in peripheral blood mononuclear cells increased by 18% one hour after the end of the stress.
A study in 2010 found that there was "significantly greater" telomerase activity in participants than controls after a three-month meditation retreat.
Telomerase deficiency has been linked to diabetes mellitus and impaired insulin secretion in mice, due to loss of pancreatic insulin-producing cells.
Mutations in TERT have been implicated in predisposing patients to aplastic anemia, a disorder in which the bone marrow fails to produce blood cells, in 2005.
Cri du chat syndrome (CdCS) is a complex disorder involving the loss of the distal portion of the short arm of chromosome 5. TERT is located in the deleted region, and loss of one copy of TERT has been suggested as a cause or contributing factor of this disease.
Dyskeratosis congenita (DC) is a disease of the bone marrow that can be caused by some mutations in the telomerase subunits. In the DC cases, about 35% cases are X-linked-recessive on the DKC1 locus and 5% cases are autosomal dominant on the TERT and TERC loci.
Patients with DC have severe bone marrow failure manifesting as abnormal skin pigmentation,leucoplakia (a white thickening of the oral mucosa) and nail dystrophy, as well as a variety of other symptoms. Individuals with either TERC or DKC1 mutations have shorter telomeres and defective telomerase activity in vitro versus other individuals of the same age
In one family autosomal dominant DC was linked to a heterozygous TERT mutation. These patients also exhibited an increased rate of telomere-shortening, and genetic anticipation (i.e., the DC phenotype worsened with each generation).
Shelterin(also called telosome) is a protein complex known to protect telomeres in many eukaryotes from DNA repair mechanisms, as well as to regulate telomerase activity. In mammals and other vertebrates, telomeric DNA consists of repeating double-stranded 5'-TTAGGG-3' (G-strand) sequences (2-15 kilobases in humans) along with the 3'-AATCCC-5' (C-strand) complement, ending with a 50-400 nucleotide 3' (G-strand) overhang. Much of the final double-stranded portion of the telomere forms a T-loop (Telomere-loop) that is invaded by the 3' (G-strand) overhang to form a small D-loop (Displacement-loop).
Shelterin co-ordinates the T-loop formation of telomeres
Shelterin has six subunits: TRF1, TRF2, POT1, RAP1, TIN2, and TPP1. They can operate in smaller subsets to regulate the length of or to protect telomeres.
TRF1 (Telomere Repeat binding Factor 1): TRF1 is a homodimeric protein that binds to the double-stranded TTAGGG region of the telomere. TRF1 along with TRF2 normally prevents telomerase from adding more telomere units to telomeres. But when telomere lengthening is required, TRF1 recruits helicases and interacts with tankyrases to facilitate the process. TRF1 is highly expressed in stem cells, and is essential for generation of induced pluripotent stem cells. TRF1 is upregulated in the brain cancer glioblastoma multiforme (GBM) in humans and mice, because of stem-cell quality of the cancer. Genetic ablation and chemical inhibition of TRF1 in mouse models of the brain cancer glioblastoma, and chemical inhibition of cultured human GBM cells inhibited tumor growth. TRF1 levels decrease with aging in humans and in mice. Increasing TRF1 in mice by gene therapy (AAV9 delivery) improved memory and other measures of health span. Conversely, inhibition of the PI3K/AKT pathway decreases TRF1, resulting in telomere-induced DNA damage. TRF1 may recruit PINX1 to inhibit telomere elongation by telomerase.
TRF2 (Telomere Repeat binding Factor 2) TRF2 is structurally related to TRF1, and helps to form T-loops. TRF2 is a homodimeric protein that binds to the double-stranded TTAGGG region of the telomere and prevents the recognition of double-strand DNA breaks. Overexpression of TRF2 leads to telomere shortening. Loss of TRF2 which leads to loss of the T-loop can activate p53 or ATM-mediated apoptosis.
Both TRF1 and TRF2 recruit the other four subunits to the telomere. Both TRF1 and TRF2 participate in telomere replication as well as in the prevention of replication fork stalling. Exercise has been shown to upregulate both TRF1 and TRF2 in leukocytes as well as endothelial cells, thereby protecting against apoptosis.
RAP1 (Repressor / Activator Protein 1): RAP1 is a stabilizing protein associated with TRF2. RAP1 inhibits DNA repair.
POT1 (Protection of Telomere 1): POT1 contains OB-folds (oligonucleotide/oligosaccharide binding) that bind POT1 to single-stranded DNA, which increase its affinity for single-stranded TTAGGG region of telomeric DNA. POT1 helps form the telomere-stabilizing D-loop. POT1 prevents the degradation of this single stranded DNA by nucleases and shelters the 3' G-overhang. POT1 suppresses ATR-mediated DNA repair. Humans only have a single POT1, whereas mice have POT1a and POT1b. POT1a inhibits DNA damage repair at the telomere, whereas POT1b regulates the length of telomeric single-stranded DNA.
TPP1 (ACD (gene)): TPP1 is a protein associated with POT1. The loss of TPP1 leads to impaired POT1 function. When telomeres are to be lengthened, TPP1 is a central factor in recruiting telomerase to telomeres. TPP1 promotes telomerase processivity in the presence of POT1. But interaction with the CST Complex limits excessive telomere elongation by telomerase. The gene which encodes for TPP1 (ACD) is distinct from the unrelated TPP1 gene on chromosome 11, which encodes tripeptidyl-peptidase I.
TIN2 (TRF1- and TRF2-Interacting Nuclear Protein 2) TIN2 is a stabilizing protein that binds to the TRF1, TRF2, and the TPP1-POT1 complex. thereby bridging units attached to double-stranded DNA and units attached to single-stranded DNA.
There are two main DNA-damage-signaling pathways that shelterin represses: the ATR kinase pathway, blocked by POT1, and the ATM kinase pathway, blocked by TRF2. In the ATR kinase pathway, ATR and ATRIP sense the presence of single-stranded DNA and induce a phosphorylation cascade that leads to cell cycle arrest. To prevent this signal, POT1 "shelters" the single-stranded region of telomeric DNA. The ATM kinase pathway, which starts from ATM and other proteins sensing double strand breaks, similarly ends with cell cycle arrest. TRF2 may also hide the ends of telomeres, just as POT1 hides the single-stranded regions. Another theory proposes the blocking of the signal downstream. This will lead to a dynamic instability of the cells over time.
The structure of the t-loop may prevent NHEJ. For NHEJ to occur, the Ku heterodimer must be able to bind to the ends of the chromosome. Another theory offers the mechanism proposed earlier: TRF2 hides the ends of telomeres.
At least four factors contribute to telomere maintenance in most eukaryotes: telomerase, shelterin, TERRA and the CST Complex. Fission yeast (Schizosaccharomyces pombe) has a shelterin complex for protection and maintenance of telomeres, but in budding yeast (Saccharomyces cerevisiae) this function is performed by the CST Complex. For fission yeast, Rap1 and Pot1 are conserved, but Tpz1 is an ortholog of TPP1 and Taz1 is an ortholog of TRF1 and TRF2.
Plants contain a variety of telomere-protecting proteins which can resemble either shelterin or the CST Complex.
Some chromosomes are depleted of telomeres, which can result in abnormal cell division and cancer. (L)
🔎
Chromosome Ends.
The ends of chromosomes are protected by specific DNA sequences called telomeres, visualized here in red. This image shows that some chromosomes are depleted of telomeres, which can result in abnormal cell division and cancer.
This image was originally submitted as part of the 2015 NCI Cancer Close Up project and selected for exhibit. (L)
Çizgenin üst bölümü bir hücrenin çekirdeğinde yer alan eşyazımı göstermektedir: Belirli bir gen için kodlama yapan bir DNA parçası mRNAya eşlemlenir; eşlemlenen DNAdaki bazlar — adenin (A), sitozin (C), guanin (G) ve thimin (T) — mRNAdaki bazlar ile özgül çiftler oluşturur. Ama DNAdaki adenin (A) mRNAdaki urasil (U) ile eşlenir. Çizgede mRNAda bulunan ve "kodon" denilen 3 bazlık bir dizi de gösterilmiştir. Eşyazımdan sonra mRNA genetik bilgileri DNAdan sitoplazmaya taşır.
Çizgenin alt bölümü bir hücrenin sitoplazmasında yer alan çeviriyi göstermektedir: mRNA ribozoma eklenir ve içinden geçer; tRNA amino asidi ribozoma taşır ve orada amino asit mRNAnın karşılık düşen bir kodon dizisine bağlanarak büyüyen bir protein zincir oluşturmak üzere önceden işlemden geçen amino asit zincirine eklenir. Tamamlanan protein daha sonra ribozomdan salınır.
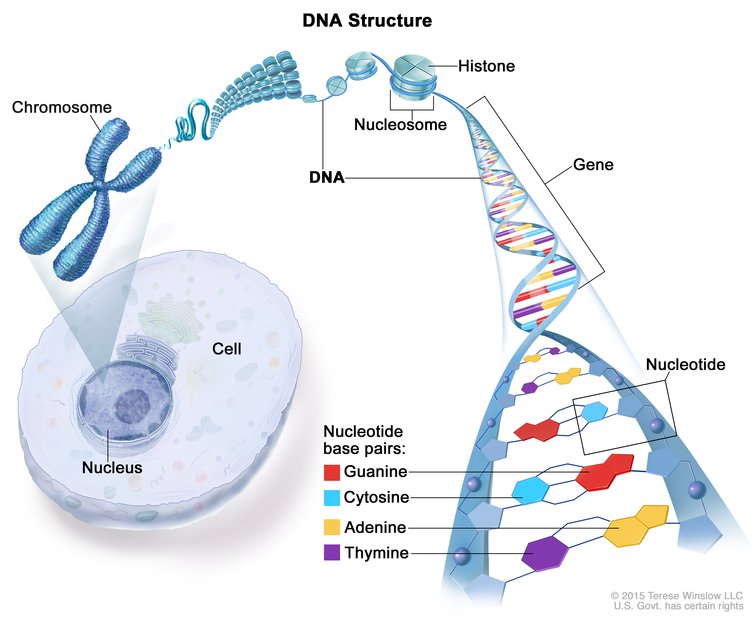
Çizge bir kromozom, nükleozom, histon, gen, ve nükleotid baz çiftlerini (guanin, sitozin, adenin ve thimin) göstermektedir. Ayrıca bir hücre ve çekirdeği de gösterilmiştir.
DNAnın yapısı. DNAnın çoğu bir hücrenin çekirdeğinin içinde bulunur ve orada kromozomları oluşturur. Kromozomlar DNAya bağlanan ve histon denilen proteinler kapsar. DNAnın iki teli vardır ve bunlar "sarmal" denilen bir spiral merdiven şekli oluşturmak üzere bükülürler. DNA nükleotidler denilen dört yapı taşından oluşur: Adenin (A), thimin (T), guanin (G) ve sitozin (C). Nükleotidler birbirine bağlanarak (A T ile, ve G C ile). baz çiftleri denilen kimyasal bağlar oluşturur. Bu çiftler iki DNA telini birbirine bağlar. Genler özgül genetik bilgiler taşıyan kısa DNA parçalarıdır.


 protrudes as a single stranded extension of the telomere.png)


.png)